


Department of Dermatology and Cutaneous Surgery, University of Miami, Miami Itch Center, Miami, FL, USA
The Janus kinase 1/3 inhibitor tofacitinib has demonstrated an antipruritic effect in two phase ΙΙΙ studies in psoriasis. However, the mechanisms behind this antipruritic effect are still unknown. We presently investigated whether tofacitinib affects spontaneous itch as well as expression of itch-related cytokines and epidermal nerve fiber density (ENFD) in the imiquimod-induced mouse model of psoriasis. Psoriasis-like skin lesions were produced by daily topical application of imiquimod to the back skin. Imiquimod treatment resulted in spontaneous scratching, which was significantly inhibited by tofacitinib treatment. Imiquimod treatment significantly increased mRNA expression of Il22, Il23, and Il31, reduced peptidergic ENFD, and increased nonpeptidergic ENFD compared to naive mice. Tofacitinib significantly decreased the expression of those cytokines and increased peptidergic ENFD with-out a significant effect on nonpeptidergic ENFD. Tofacitinib may inhibit psoriatic itch through inhibition of cytokine expression as well as modulation of epidermal innervation.
Key words: JAK inhibitor; chronic itch; psoriasis; scratching; IL-31.
Accepted Nov 20, 2018; E-published Nov 21, 2018
Acta Derm Venereol 2019; 99:
Corr: Tasuku Akiyama, PhD, Department of Dermatology and Cutaneous Surgery, University of Miami, Miami Itch Center, 1600 NW 10th Ave RMSB2063, Miami, FL 33136, USA. E-mail: takiyama@miami.edu
Tofacitinib, a drug that inhibits Janus kinases 1 and 3, has been shown to reduce itch in clinical trials of psoriasis. However, the exact mechanism for this anti-itch effect is still unclear. This study investigates the effects of tofacitinib treatment in a mouse model of psoriasis. Tofacitinib reduced scratching in this model. Additionally, tofacitinib rescued the increased mRNA levels of Il22, Il23, and Il31, 3 cytokines that are known to be related to itch. Finally, tofacitinib recovered the reduction in peptidergic nerves in the epidermis of psoriasis model mice. Therefore, tofacitinib could reduce itch through regulation of cytokines and epidermal nerves.
Tofacitinib is a targeted small molecule that preferentially inhibits Janus kinase (JAK)-1/3. This inhibition of JAK-1/3 blocks γ-chain cytokines, including interleukin (IL)-2, IL-4, IL-7, IL-9, IL-15, and IL-21, that could contribute to skin inflammation (1). Tofacitinib demonstrated a significant antipruritic effect in two phase ΙΙΙ studies of patients with moderate to severe plaque psoriasis and one phase ΙΙa study of patients with mild to moderate atopic dermatitis (2–4). Interestingly, tofacitinib has a rapid antipruritic effect which is independent from improvements in other clinical signs of psoriasis, such as erythema, induration, and scaling (5). A recent study suggests that tofacitinib acts directly on the transient receptor potential vanilloid 1 ion channel expressed in sensory neurons to exert antipruritic effects (6). This could explain the rapid onset of improvement in pruritus. However, the mechanism behind the antipruritic effects of tofacitinib has not been fully understood.
Psoriasis is an inflammatory skin disease that is driven by Th1 and Th17 cells (7). IL-17A, IL-17F, IL-21, and IL-22 are found at increased levels in psoriatic skin (8, 9). Interestingly, RNA-seq analysis demonstrated that mRNA transcripts for Il17a, Il22, Il23, and Il31 were elevated in the itchy skin of both atopic dermatitis and psoriasis patients compared to non-itchy skin (10). Knockout mouse studies have revealed the role of cytokines (e.g. IL-31, thymic stromal lymphopoietin (TSLP), IL-4, IL-33, and tumor necrosis factor-α) in itch (11–15). Particularly, IL-31 and TSLP directly act on their cognate cytokine receptors expressed by primary sensory neurons and keratinocytes, respectively, to elicit itch.
Dynamic changes in epidermal innervation have been observed under chronic itch conditions. Epidermal hyper-innervation by itch-signaling fibers is considered to be one cause of chronic itch (16), and increased epidermal nerve density has been frequently reported in humans with chronic itch conditions including psoriasis (16–20). In line with this, intraepidermal nonpeptidergic nerve density is increased in animal models of dry skin itch and psoriatic itch (21–23). In contrast, conflicting results have been reported for epidermal nerve density in human with chronic itch. Reduced epidermal nerve density has been reported in patients with psoriatic itch as well as prurigo nodularis (17, 24, 25). Additionally, intraepidermal peptidergic nerve density is decreased in animal models of psoriatic itch (23). In these cases, reduced primary afferent drive toward itch-inhibitory interneurons could contribute to disinhibition of itch.
We presently investigated whether tofacitinib inhibited spontaneous itch in the imiquimod-induced mouse model of psoriasis. In addition, we tested if tofacitinib prevented an increase in itch-related cytokines and dynamic changes in epidermal innervation in this model.
Experiments were performed using adult male C57BL/6J mice (24–29 g) under a protocol approved by the University of Miami Institutional Animal Care and Use Committee (Protocol #17-008).
Fur on the rostral back was trimmed with electric clippers and then removed with an electric shaver. Each mouse received a daily topical application of 62.5 mg Aldara cream (5% imiquimod, Meda Pharmaceuticals, Somerset, NJ) on the shaved back skin (2.5 × 2 cm) for 7 consecutive days. Naive mice were shaved but did not receive any imiquimod treatment.
Tofacitinib (LC Laboratories, Woburn, MA) was dissolved in a sterile solution of 50% DMSO, 10% PEG 300, and 40% water. Under isoflurane anesthesia, ALZET osmotic mini pumps (Durect Corp, Cupertino, CA) were implanted in the peritoneal cavity of imiquimod-treated mice to deliver a dose of 15 mg/kg/day tofacitinib or vehicle beginning immediately after the implantation. Mini pumps were implanted 7 days before the start of imiquimod treatment. As such, there were 3 experimental groups: control mice with no imiquimod treatment (“naive”), mice that received topical imiquimod treatment and systemic vehicle treatment (IMQ+Veh), and mice that received topical imiquimod treatment and systemic tofacitinib treatment (IMQ+Tof).
Mice were habituated twice to a Plexiglas recording arena for 60 min before testing. At baseline (Day 0) and 20 to 22 h after each topical application, mice were videotaped from above for 60 min. The number of videotaped scratch bouts was counted by a trained observer blinded to the treatment condition. A scratch bout was defined as one or more rapid back-and-forth hind paw motions directed toward and contacting the treated area and ending with licking or biting of the toes or placement of the hind paw on the floor. Hind paw movements directed away from the treated area (e.g., ear-scratching) and grooming movements were not counted (26–28).
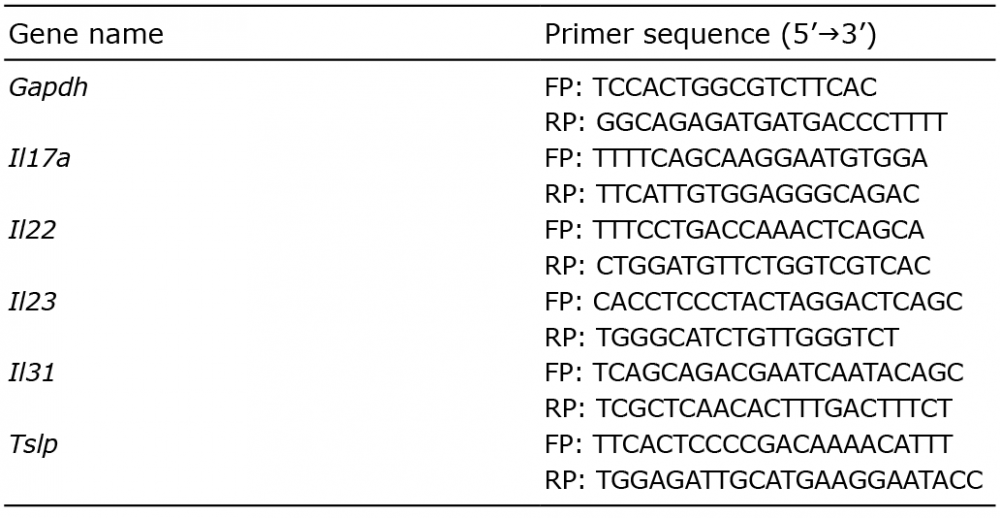
Animals were euthanized under sodium pentobarbital anesthesia, and skin samples were immediately collected, preserved in RNAlater (Qiagen, Valencia, CA), and stored at –80°C. Total RNA was extracted using Direct-zol RNA Mini Prep (Zymo Research, Irvine, CA). Reverse transcription of 0.5 μg total RNA was performed using ProtoScript® II First Strand cDNA Synthesis Kit (New England Biolabs, Ipswich, MA). Amplification of GAPDH cDNA was used for normalization. Real-time RT-PCR was performed using Fast Plus EvaGreen qPCR Master Mix (Biotium, Hayward, CA) on a QuantStudio 3 Real-Time PCR System (Thermo Fisher Scientific Inc., Waltham, MA). Forty cycles of amplification were performed involving sequential denaturation at 95°C for 15 s and annealing/extension at 55–56°C for 30 s. Assays were validated using serial dilutions and confirmation of equal amplification efficiencies of the cDNA of interest and the GAPDH cDNA. Fold differences in expression were calculated using the comparative ΔCt method by standardizing against GAPDH expression. All primer pairs are listed in Table I.
Table I. List of primers; forward primer (FP) and reverse primer (RP)
Animals were euthanized under sodium pentobarbital anesthesia, and the skin was immediately dissected. Skin was fixed in Zamboni fixative solution (Newcomer Supply, Middleton, WI) followed by 30% sucrose, frozen in optimal cutting temperature (OCT) compound (Tissue-Tek, Sakura Finetek, Torrance, CA), and cut in 40-µm sections on a cryostat. The sections were incubated with 5% donkey serum and 0.2% Triton X-100 in PBS and then immunostained with either rabbit CGRP (1:300; Peninsula Laboratories International Inc., San Carlos, CA), P2X3 (1:200; NeuromicsInc., Edina, MN), or PGP9.5 antibody (1:200; EMD Millipore, Burlington, MA) at 4°C overnight, followed by incubation with the corresponding secondary antibody conjugated with AlexaFluor 488 (1:300; Life Technologies Inc., Grand Island, NY) or AlexaFluor 594 (1:300; Life Technologies Inc.) for 2 h. All sections were counterstained with 4’,6-diamino-2-phenylindole (DAPI) in the mounting medium (Vector Laboratories, Burlingame, CA). Images were captured from 10–12 skin sections from each animal at 20X magnification (4–6 mice per group) and evaluated by a trained observer blinded to the treatment condition. The number of labeled nerve fibers crossing the dermal–epidermal junction (DEJ) was counted. The length of the DEJ (in mm) was measured using ImageJ. Epidermal nerve fiber density (ENFD) was calculated as the number of epidermal nerve fibers divided by the length of the DEJ.
Between-group comparisons were made by one-way ANOVA followed by Tukey post hoc test or two-way RM ANOVA followed by Bonferroni post hoc test. In all cases p < 0.05 was considered to be significant.
To test the effects of tofacitinib on psoriatic itch, we used the imiquimod-induced psoriasis mouse model (Fig. 1A). Tofacitinib significantly reduced counts of spontaneous scratch bouts on Day 7 in imiquimod-treated mice (Fig. 1B). Of note, we did not observe a recovery in skin inflammation after treatment with the tofacitinib (data not shown).
Fig. 1. Effects of tofacitinib on spontaneous itch in psoriasis model. A) Timeline for imiquimod-induced psoriasis-like skin inflammation model and implantation of osmotic mini pumps. Osmotic pumps were implanted 7 days before the first imiquimod treatment and delivered either vehicle (50% DMSO, 10% PEG 300, 40% distilled water) or tofacitinib. Mice were treated with imiquimod every day for 7 days. Behavior testing and/or skin dissection were conducted 20–22 h post-treatment. B) Time-dependent changes in scratch bouts in imiquimod-treated mice with or without tofacitinib implantation. Spontaneous scratching was measured on pretreatment day 0 and imiquimod treatment days 1, 3, 5, and 7. Black dots (?) and white squares (?) show, respectively, tofacitinib- and vehicle-treated groups. Error bars are SEM (n = 8). *p < 0.05, F(4,56)=24.1, significant difference from vehicle-treated group, two-way RM-ANOVA followed by Bonferroni test.
Th17 and itch-associated cytokine mRNA levels were measured in skin from naive, IMQ+Veh, and IMQ+Tof mice. mRNA expression levels of Il17a, Il22, Il23, and Il31 were elevated in imiquimod-treated skin compared to naive skin (Fig. 2A–D). Levels of Il22, Il23, and Il31 were significantly decreased by treatment with tofacitinib. While Tslp mRNA was not changed in the imiquimod model compared to naive skin, Tslp mRNA was significantly elevated in IMQ+Tof mice (Fig. 2E).
Fig. 2. Effects of tofacitinib on cytokine expression in psoriasis model. Levels of Il17a (A), Il22 (B), Il23 (C), Il31 (D), and Tslp (E) in the skin were measured using RT-qPCR. Error bars are SEM (n = 5–8). *p < 0.05, significant difference from naive mice, #p< 0.05, significant difference from IMQ+Veh-treated group, one-way ANOVA followed by Tukey test.
Total ENFD was decreased in the IMQ+Veh group compared to naive control (naive: 71.2 ± 7.4 nerve fiber counts/mm epidermis; IMQ+Veh: 58.3 ± 4.3 counts/mm) (Fig. 3A, B, D). Total ENFD was decreased in the IMQ+Tof treated group compared to the naive control and the IMQ+Veh group (IMQ+Tof: 51.4 ± 2.8 counts/mm) (Fig. 3A–D). However, the changes in total ENFD were not statistically significant. CGRP-positive peptidergic ENFD was significantly decreased in the IMQ+Veh group compared to the naive control (Fig. 3E, F, H). This decrease was significantly blocked by treatment with tofacitinib (Fig. 3F–H). P2X3-positive nonpeptidergic ENFD was significantly increased in the IMQ+Veh group compared to the naive control (Fig. 3 I, J, L). Treatment with tofacitinib did not prevent this increase (Fig. 3J–L).
Fig. 3. Effects of tofacitinib on epidermal nerve fiber density in psoriasis model. (A–L) Skin was dissected from mice left untreated (naive, A, D, E, H, I, L) or after 7 days of imiquimod (IMQ) treatment with systemic vehicle (B, D, F, H, J, L) or tofacitinib (C, D, G, H, K, L). (A–C) Skin sections were immunostained with an antibody for PGP9.5 to visualize all nerve fibers. White arrows indicate nerve fibers crossing the dermal–epidermal junction. Scale bar indicates 50 µm. (D) Total epidermal nerve fiber density (ENFD) was quantified in naive mice (grey bars) and IMQ+Veh– (white bars) and IMQ+Tof-treated mice (black bars). Error bars are SEM. n = 4/group. (E–G) Skin sections were immunostained with an antibody for CGRP, a marker for peptidergic nerves. (H) As in D for CGRP+ nerve fibers. *p < 0.05, significant difference from naive mice, #p < 0.05, significant difference from IMQ+Veh–treated group, one-way ANOVA followed by Tukey test, n = 4/group. (I–K) Skin sections were immunostained with an antibody for P2X3, a marker for nonpeptidergic nerves. (L) As in D for P2X3+ nerve fibers. *p < 0.05, significant difference from naive mice (one-way ANOVA followed by Tukey test, n = 4/group).
In the present study, sustained systemic delivery of tofacitinib inhibited spontaneous itch-related behaviors in the imiquimod-induced psoriasis model. We investigated whether this antipruritic effect was associated with suppression of itch-related cytokines and/or alterations in epidermal nerves. We found that tofacitinib prevented the increased mRNA expression of cytokines, including Il22, Il23, and Il31, that are elevated in the imiquimod-induced model of psoriasis and also in the itchy skin of humans with atopic dermatitis and psoriasis (10). Our result confirms similar findings reported by other groups. Topical applications of tofacitinib inhibited the elevated expression of IL-22 in imiquimod-treated mouse ear skin and IL-31 in toluene-2,4-diisocyanate-treated mice (an allergic dermatitis model) (29, 30). In line with this, in vitro studies demonstrated that tofacitinib inhibited IL-23 expression in dendritic cells and IL-22 expression in Th17 cells as well as Th-cell differentiation of CD4+ T cells into Th2 cells that can release IL-31 (31). IL-31 has been identified as a major itch-related cytokine in several pruritic diseases (10, 32–36). If tofacitinib exerts its antipruritic effect via suppression of IL-31, it may be a useful anti-itch therapy for a variety of chronic itch conditions including atopic eczema, contact dermatitis, lichen amyloidosis and cutaneous T-cell lymphoma.
JAK is the downstream signal of TSLP and the upstream signal of IL-23 (37, 38). Here, we showed that tofacitinib elevated the mRNA expression of Tslp and decreased the mRNA expression of Il-23 in imiquimod-treated mice. Similarly, vitamin D3 treatment upregulated TSLP and downregulated IL-23 in psoriasis patients, suggesting that TSLP and IL-23 are inversely regulated during the amelioration of psoriasis (39). TSLP can upregulate IL-17A through CXCR2 expressed by keratinocytes (40). Additionally, IL-17A can elevate the expression levels of IL-33, which can potentiate the induction of TSLP, in keratinocytes (41, 42). As such, there is likely feedback upregulation of TSLP through the production of IL-17A and IL-33 in keratinocytes. Our study showed that tofacitinib did not reduce the elevated mRNA expression of Il17a. Similarly, topical application of tofacitinib did not significantly inhibit Il17a mRNA expression of the ear skin of imiquimod-treated mice (30). While tofacitinib decreased the expression of IL-23, the upstream activator of IL-17A, the TSLP feedback upregulation loop may have compensated for the decreased production of IL17A.
One possible limitation of the present study is that we used naive mice rather than vehicle cream-treated mice as a control group for the imiquimod-treated groups. While it has been reported that vehicle cream may induce inflammation (43), we did not observe scratching behavior or touch-evoked itch in mice treated with vehicle cream for 7 days (44), suggesting that vehicle treatment alone does not significantly upregulate itch signaling pathways.
Consistent with our previous study (23), the density of nonpeptidergic epidermal nerves was significantly increased in the imiquimod-treated mice compared to naive mice. We have previously reported that the growth factor neurturin contributes to an increase in the epidermal density of nonpeptidergic nerves in this model, and this increase is a factor for chronic itch in psoriasis (23). However, tofacitinib did not recover the non-peptidergic ENFD, making it unlikely that tofacitinib exerts its antipruritic effect through blocking this mechanism.
As we reported previously (23), the density of peptidergic epidermal nerves was significantly decreased in the imiquimod-treated mice compared to naive mice. Tofacitinib significantly increased the peptidergic ENFD. Peripheral input is necessary to activate itch-inhibitory systems, such as B5-I spinal interneurons (45). The re-innervation of peptidergic epidermal nerves may rescue the inhibition of itch by scratching and thus contribute to the antipruritic effects of tofacitinib. This finding suggests a novel mechanism for reduction of itch by tofacitinib and points to the potential of peptidergic ENFD to be an important target for treatment of chronic itch.
This work was supported by Pfizer grant (W1203521: GY and TA) and National Institutes of Health grant (AR063228: TA). TH. has received scholarships from the Uehara Memorial Foundation and the Japanese Society of Allergology. KMS was supported by the Lois Pope LIFE Fellows Program.
Conflict of interest: GY has served in advisory board and consultant for Pfizer, Novartis, Eli Lilly, Sanofi, Galderma, Trevi, Menlo, Sienna, UCB.


 Click to show fullsize
Click to show fullsize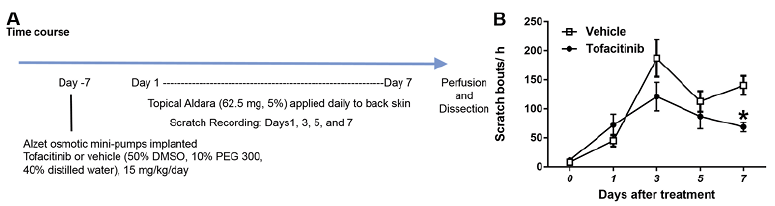 Click to show fullsize
Click to show fullsize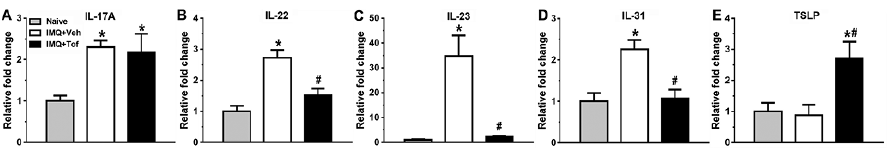 Click to show fullsize
Click to show fullsize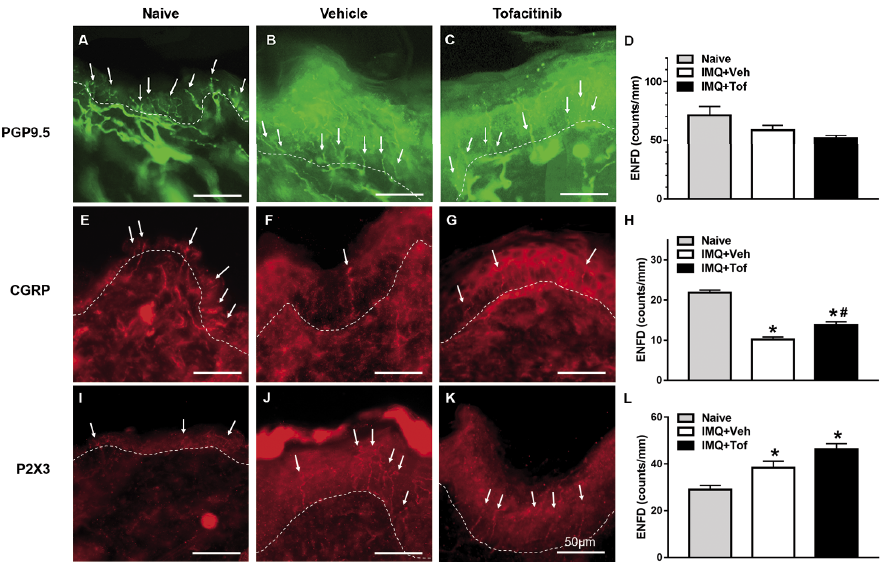 Click to show fullsize
Click to show fullsize